
m6A RNA甲基化专题
m6A RNA甲基化
一、什么是RNA甲基化
表观基因组指的是修饰DNA及其蛋白支架的一系列化学标记可以用来帮助解释拥有相同DNA的细胞会发育成众多的专门类型,构成不同的组织。大部分表观遗传研究都集中在与DNA以及DNA包裹的组蛋白相关的标记上。
1974年首次在mRNA中发现腺嘌呤上的一个甲基,这个被修饰的碱基被称作N6-甲基腺苷(N6-methyladenosine),简写为m6A。2012年,两支团队独立发布了m6A的首个位点图谱,这些研究揭示了来自约7000个基因的mRNA上的12000多个甲基化位点。
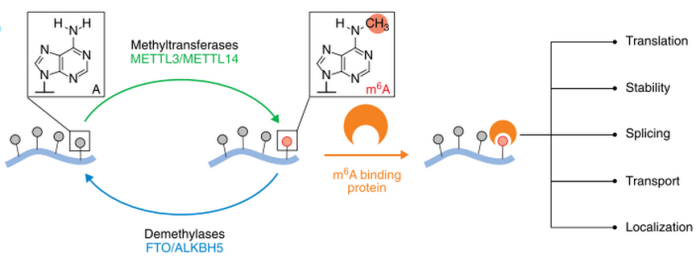
在过去几年中,一系列重量级的文章的出现,揭示了调控这些标记所涉及的一些机制。RNA甲基化中的m6A首先在甲基化转移酶的作用下对RNA上的腺嘌呤(A)进行m6A修饰,再在去甲基化酶的作用下对已发生m6A修饰的RNA进行去甲基化修饰。最后发生m6A修饰的RNA通过被甲基化阅读蛋白识别后,行使下游的一系列功能,包括miRNA加工、mRNA出核翻译及剪切等。每个标记都需要一个“写入器(writer)”来放置,一个“擦除器(eraser)”消除,还需要“阅读器(reader)”读取。随着这些蛋白质的身份浮出水面,我们了解到m6A不仅会影响RNA剪接,还会影响翻译和RNA稳定性。

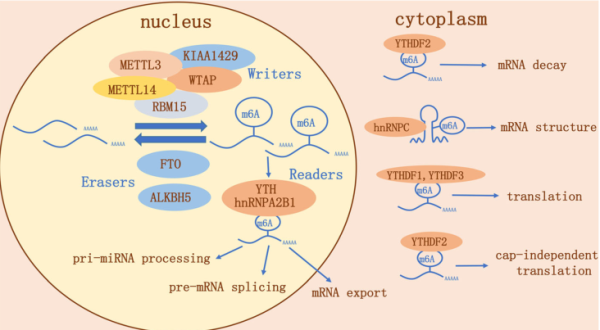
M6a基因调控途径在肿瘤里的研究目前处于起步阶段,根据文献整理,目前的研究涉及8种肿瘤以及6个M6a组成分子[1]。
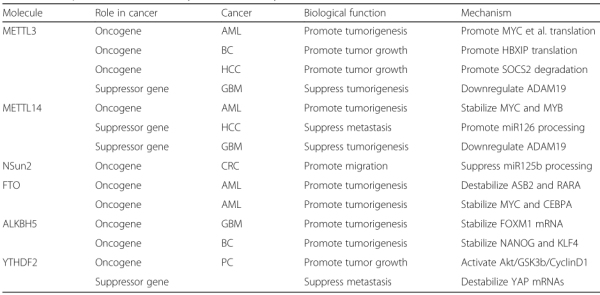
比如,Li等通过基因表达芯片,发现m6A去甲基化酶FOT在特定的急性髓系白血病中表达量升高后,构建FOT的敲除的白血病细胞系,体内体外实验发现细胞生长抑制。结合m6A-seq技术,发现在白血病细胞系中,FOT的靶基因为ASB2和RARA[2]。采用类似思路,Zhang等在胶质母细胞瘤样干细胞中发现去甲基化酶ALKBH5通过促进FOXM1表达进而保证干细胞的增值及成瘤能力[3]。Su等发现代谢物R-2HG通过抑制FTO活性,进而促进白血病细胞中m6A的发生水平,以此降低了MYC/CEBPA转录物的含量而表现出抗肿瘤活性[4]。

二、m6A甲基化研究方法
(1)寻找m6A相关酶在骨关节炎中的异常表达情况。鉴于mRNA的测序和芯片数据较全,建议从数据库中寻找,比如cbioportal, oncomime和GEO datasets等。
(2)细胞及动物的表型实验。通过敲低和过表达该m6A修饰酶,检测细胞的变化,包括细胞增殖,凋亡,验证变化等指标。
(3)通过m6A-seq和RNA-seq的结合,寻找m6A含量变化和RNA分子数量变化最大的mRNA。
1、开启测序前的一些“预实验”
该步所有实验为可选项,也可以跳过这部分直接进入测序部分,来挖掘哪些基因在甲基化修饰上产生了差异。
(1)找到差异writers、erasers和readers或者检测整体m6A水平
首先,要找到与RNA甲基化相关的酶是否有差异表达。既可以是药物处理前VS药物处理后,也可以是肿瘤组织VS癌旁组织,甚至是发育的时间顺序,非模式生物不同的组织器官做比较也是考虑的研究方向。
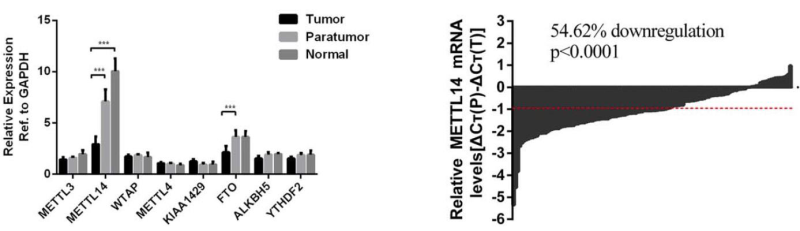
(Ma J et al. (2017) Hepatology, 62(2): 529)
如上图所示,作者对肝癌组织、癌旁组织以及正常组织中几种常见的酶都进行了qPCR验证,发现FTO和METTL14有差异表达。
另外通过GWAS以及各种人群大队列分析得到的易感基因正好和RNA甲基化有关,那么这项课题开展下去的意义会更大。这时候RNA甲基化被赋予了新的生命,是上游课题的延续。即全基因组重测序/全外显子测序/靶向捕获测序→SNP和Indel分析→GWAS→易感基因→细胞实验/动物实验→下游通路和分子机制→最终表型。
(Li et al. (2017) Cancer Cell, 31(1):127-141)
另一种方式就是从已发表的芯片数据中,直接挖掘几种酶是否有差异表达(省钱的)。如上图Cancer Cell上的论文,前期就是从网上队列数据中筛选出FTO基因才继续进入下游的细胞实验和动物实验。
至于检测m6A整体水平,可采用LC-MS/MS或酶标仪比色法的方式进行。酶标仪相对于LC-MS/MS普及程度更高,可采用EpiQuik m6A RNA Methylation Quantification Kit的试剂盒直接进行检测,方便快捷。
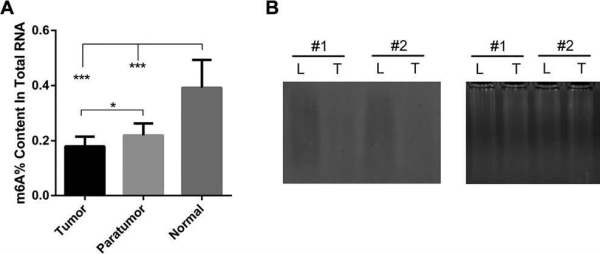
(Ma J et al. (2017) Hepatology, 62(2): 529)
如上图所示,对肝癌组织、癌旁组织和正常组织在酶标仪上进行了m6A整体的水平检测后发现,肝癌组织m6A整体水平最低,而正常组织中m6A水平最高。
此外,在细胞中对几种酶进行敲低和过表达后,也可以选择使用酶标仪来检测整体的m6A水平是否发生变化。
(2)甲基化酶敲低和过表达细胞实验
这里对几种已检测的甲基化酶进行敲低或过表达,当然这一步也可以在一开始直接进行。之后观察细胞表型和动物表型,建立起相关性关系。通常细胞实验比较容易开展,可以开始进行对应敲低/过表达体系的建立。
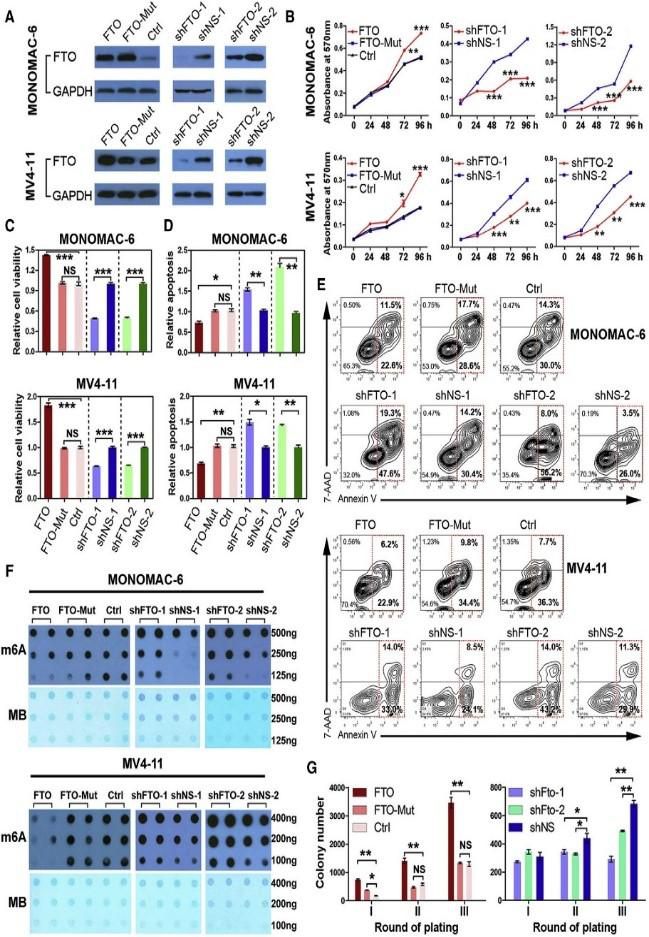
(Li et al. (2017) Cancer Cell, 31(1):127-141)
如上图所示,在急性髓细胞白血病AML中,FTO存在表达量显著上升现象。作者在AML细胞系中,对FTO这个基因进行敲低和过表达后发现AML细胞m6A整体水平降低,凋亡减少。敲低FTO趋势相反。
(3)甲基化酶敲低和过表达动物模型实验
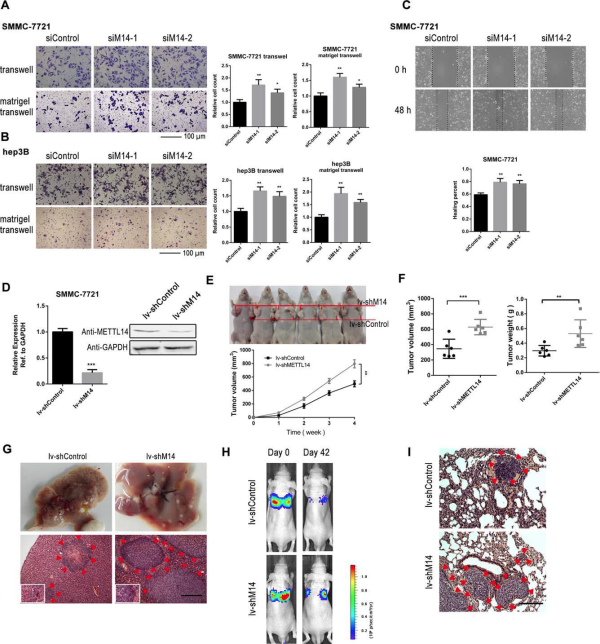
(Ma J et al. (2017) Hepatology, 62(2): 529)
动物模型通常为可选项,老师可以根据自己的经费和实验整体设计来安排。如上图所示,作者在小鼠肿瘤模型中对METTL14进行敲低后发现肝癌转移能力增强。
2、m6A-seq挖掘靶基因
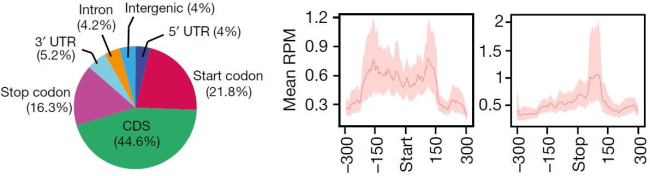
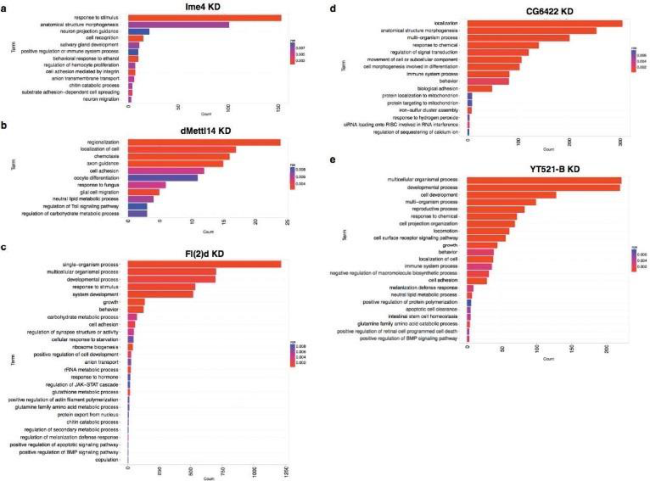
(Lence et al. (2016) Nature, 540(7632):242-247)
通过m6A-seq测序,可以知道究竟哪些基因在RNA甲基化修饰上有差异。如上图所示,作者对果蝇SR2+细胞进行m6A-seq和转录组测序后发现,差异基因都富集在神经功能上。
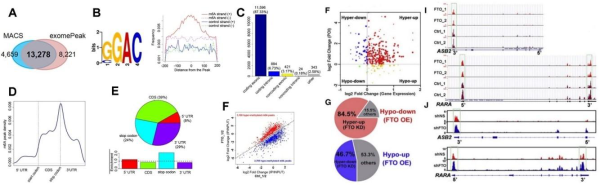
(Li et al. (2017) Cancer Cell, 31(1):127-141)
如果最后的差异基因或者存在甲基化修饰差异的基因过多怎么办?
可以结合自己先前课题的研究,在这些差异基因中寻找是否包含原有已知的通路和基因。如上图所示,通过m6A-seq作者在AML发现了几百个差异基因。初步筛选一般包括功能富集分析等方法。当几百个基因被限定到11个差异基因时,作者从中发现了RARA和ASB2这2个抑制白血病细胞生长的基因与全反式维甲酸相关。先前研究表明全反式维甲酸治疗后ASB2和RARA显著上调。这样子,在一大堆候选基因中,范围继续缩小。
3、靶基因细胞实验
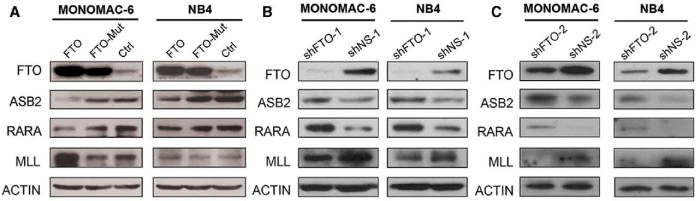
(Li et al. (2017) Cancer Cell, 31(1):127-141)
如上图所示,在确定ASB2和RARA为去甲基化酶FTO的靶基因后,除了常规的qPCR+WB验证外,作者还进行了补救实验(rescue)以及m6A-IP-qPCR(也叫MeRIP-qPCR)。作者用细胞实验对ASB2和RARA进行敲低并发现AML细胞表型与FTO过表达相似,表型被补救。
高分文章会有两个大方向——临床和基础研究。
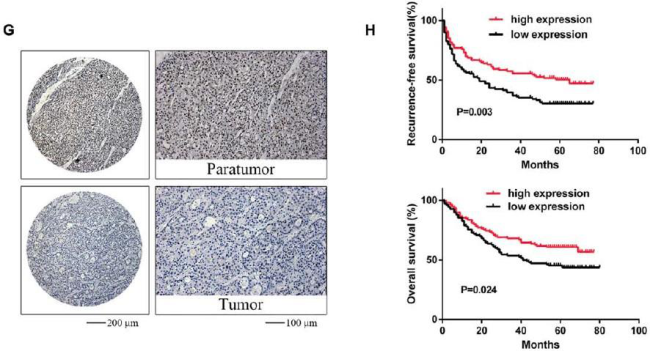
(Ma J et al. (2017) Hepatology, 62(2): 529)
在临床方面,通过上述实验挖掘到的甲基化酶或者是被调控的下游通路某一个基因或miRNA具有一定的临床意义会更加容易冲击高分文章。如上图所示,在HCC中METTL14低表达与转移侵染能力在表型上建立了相关性,METTL14在HCC中显著降低也与性别、乙肝病毒抗原以及微血管转移相关,METTL14与病人生存周期相关。所以作者最后得出结论,METTL14作为潜在的预后marker与HCC的转移相关。通过进一步分析发现,METTL14调控miR-126,而miR-126本身能够抑制肿瘤细胞的转移能力。那么在之前的基础上,挖掘到了更深层次的信息。
在基础生物学方面,可以通过Co-IP甚至是SILAC挖掘到与甲基化酶有互作的一些新蛋白并进行下游验证。或者可以购买几种酶的抗体进行RIP实验后,对富集到的RNA进行分析挖掘,找到几种酶有互作的RNA甚至是互作区域。接下来对特定的motif进行突变等实验进行验证。在这些实验完成后,如果能发现一种全新的机制一般容易冲击顶级期刊。
参考文献:
[1] Pan Y, Ma P, Liu Y, et al. Multiple functions of m 6 A RNA methylation in cancer[J]. Journal of Hematology & Oncology, 2018, 11(1):48.IF=6.35
[2] Li Z, Weng H, Su R, et al. FTO Plays an Oncogenic Role in Acute Myeloid Leukemia as a N6-Methyladenosine RNA Demethylase[J]. Cancer Cell, 2016, 31(1):127.IF=27.407
[3] Zhang S, Zhao B S, Zhou A, et al. m 6 A Demethylase ALKBH5 Maintains Tumorigenicity of Glioblastoma Stem-like Cells by Sustaining FOXM1 Expression and Cell Proliferation Program[J]. Cancer Cell, 2017, 31(4):591.IF=27.407
[4] Su R, Dong L, Li C, et al. R-2HG Exhibits Anti-tumor Activity by Targeting FTO/m6A/MYC/CEBPA Signaling[J]. Cell, 2017, 172(1-2):90-105. IF=30.4
[5] Meyer, K., & Jaffrey, S. R. (2017). Rethinking m6A Readers, Writers, and Erasers. Annual Review of Cell & Developmental Biology, 33(1), 319.
[6] Fu, Y., Dan, D., Rechavi, G., & He, C. (2014). Gene expression regulation mediated through reversible m6A RNA methylation. Nature Reviews Genetics, 15(5), 293-306.
[7] Roundtree, I. A., Evans, M. E., Pan, T., & He, C. (2017). Dynamic RNA Modifications in Gene Expression Regulation. Cell, 169(7), 1187.
